
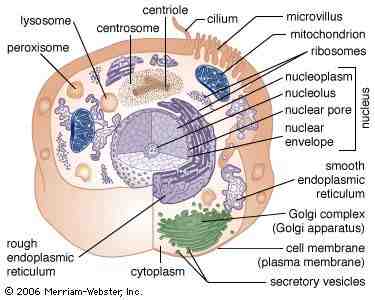
Comunicare celulară
Comunicarea intercelulară
Formarea unui organism multicelular începe cu o mică colecție de celule similare dintr-un embrion și se desfășoară prin diviziune celulară continuă și specializare pentru a produce o întreagă comunitate de celule care cooperează, fiecare cu rolul său în viața organismului. Prin cooperarea celulară, organismul devine mult mai mult decât suma părților sale componente.
etapele timpurii ale dezvoltării umane Ovulul conține o mică colecție de celule în stadiile incipiente ale dezvoltării umane. Pe măsură ce celulele se divid (A – D), acestea sunt separate în diferite regiuni ale ovulului. Fiecare regiune a ovulului transmite un set unic de semnale chimice către celulele din apropiere. Astfel, semnalele detectate de o celulă diferă de cele detectate de celulele vecine. În acest proces, cunoscut sub numele de determinare celulară, celulele sunt programate individual pentru a le direcționa spre dezvoltarea în diferite tipuri de celule. Encyclopædia Britannica, Inc.
Un ovul fertilizat se înmulțește și produce o întreagă familie de celule fiice, fiecare adoptând o structură și o funcție în funcție de poziția sa în întregul ansamblu. Toate celulele fiice conțin aceiași cromozomi și, prin urmare, aceleași informații genetice. În ciuda acestei moșteniri comune, diferite tipuri de celule se comportă diferit și au structuri diferite. Pentru ca acest lucru să fie cazul, ei trebuie să exprime diferite seturi de gene, astfel încât să producă proteine diferite în ciuda lor identice strămoși embriologici .
Semnalizare celulară
Semnalizarea celulară (cell signaling în engleză ) face parte din orice proces de comunicare care guvernează activitatea celulară de bază și coordonează toate activitățile celulare. Capacitatea celulelor de a percepe și de a răspunde corect la micro-mediul lor stă la baza dezvoltării, reparării țesuturilor, imunității și homeostaziei țesuturilor normale. Erorile în interacțiunile de semnalizare și procesarea informațiilor celulare sunt responsabile de boli precum cancerul , autoimunitatea și diabetul. Prin înțelegerea semnalizării celulare, bolile pot fi tratate mai eficient și, teoretic, se pot crea țesuturi artificiale.
Biologia sistemelor studiază structura de bază a rețelelor de semnalizare celulară și modul în care schimbările din aceste rețele pot afecta transmisia și fluxul de informații (transducția semnalului). Astfel de rețele sunt sisteme complexe în organizarea lor și pot prezenta o serie de proprietăți emergente, inclusiv bistabilitatea și ultra-sensibilitatea. Analiza rețelelor de semnalizare celulară necesită o combinație de abordări experimentale și teoretice, inclusiv dezvoltarea și analiza simulărilor și modelării.
Generalitate
Cei responsabili de comunicarea celulară sunt în general molecule extracelulare secretate de alte celule sau de celula însăși care pot acționa asupra celulei în sine, pe distanțe scurte sau pe distanțe mari. Aceste molecule sunt capturate de receptori , care sunt adesea proteine transmembranare situate în membrana plasmatică cu un loc de legare situat în spațiul extracelular. Una dintre cele mai frecvente categorii de molecule care declanșează un răspuns celular sunt hormonii . Legarea ligandului (molecula extracelulară) cu receptorul său specific declanșează activarea acestuia într-o gamă largă de moduri posibile și, la rândul său, receptorul declanșează una sau mai multe căi de semnalizare intracelulare. Căile de semnalizare implică în mare parte proteine de semnalizare, ele pot fi relativ simple, dar mai des, în organismele superioare, sunt complexe și interconectate între ele, nu au direcționalitate specifică și pot fi citite atât din membrana plasmatică către citoplasmă sau nucleu, cât și viceversa.
Cu toate acestea, proteinele nu sunt singurii actori ai acestor căi de semnalizare care implică deseori și ioni anorganici, fosfolipide, steroizi și derivații lor. În cele din urmă, aceste proteine converg semnalizarea în proteine efectoare, a căror conformație este modificată de reacții chimice adecvate care pot duce la activarea sau inactivarea lor. Aceste proteine includ, de exemplu, canale ionice , proteine de reglare a genelor, enzime , proteine structurale ale citoscheletului, receptori intracelulari. Aceste proteine acționează asupra altor ținte declanșând răspunsul final la acea cale de semnalizare dată, care poate fi alterarea expresiei genelor, alterarea metabolismului uneia sau mai multor substanțe, crearea de complexe proteice, mișcarea celulei prin acțiune. citoscheletul.
Proteinele intracelulare au nevoie de un timp de înjumătățire scurt și de o rată de rotație rapidă pentru a fi reglementate de celulă, de fapt, dacă timpul de înjumătățire ar fi ridicat, intervalul posibil de reglare ar fi mai mic. Cifra de afaceri ajută și la modificarea duratei răspunsului, deoarece adesea aceste proteine sunt modificate (de exemplu prin fosforilare) pentru a fi activate, rezultând că atunci când o proteină activată este degradată semnalul devine din ce în ce mai slab pe măsură ce este înlocuit cu o proteină. gata să restabilească alerta în urma modificării corespunzătoare.
Molecule de semnalizare extracelulară
Moleculele de semnal cunoscute sunt mii și includ ioni anorganici, gaze, aminoacizi, peptide, proteine, nucleotide, steroizi, lipide, derivați ai acizilor grași. În general, acestea sunt secretate de o celulă de semnalizare pentru a transmite un semnal către o celulă țintă care se află la o distanță variabilă. Ele pot fi secretate în matricea extracelulară sau în lichide specifice, dintre care cel mai frecvent este sângele. Concentrația moleculelor de semnalizare este de obicei foarte scăzută ( 10-8 M sau mai mică) și receptorii pe care aceștia se leagă au o afinitate foarte mare pentru acești liganzi (K la 10 -8 L mol sau mai mare), deci sunt foarte specifici. Fiecare celulă primește anumite tipuri de molecule de semnalizare pe baza receptorilor pe care îi exprimă la suprafață sau în citoplasmă. Specializarea unei celule determină ce tipuri de receptori sunt sintetizați, ce tipuri de proteine de semnalizare intracelulare și efectoare și cum acționează moleculele de semnalizare asupra lor.
Prin urmare, două celule diferite care primesc aceeași moleculă de semnal pot răspunde diferit. Diversitatea răspunsului celular poate depinde și de factori cantitativi. Unele celule, de exemplu, răspund diferit în funcție de concentrația mai mult sau mai puțin mare a unei molecule de semnalizare date, uneori dând naștere unor efecte opuse și aparent paradoxale. Concentrația moleculei de semnalizare joacă un rol fundamental în dezvoltarea unui organism în care grupurile de celule nediferențiate sau parțial nediferențiate dobândesc destine celulare diferite, un număr diferit de receptori, proteine de semnalizare și efectoare pentru molecula respectivă și o specializare diferită în funcție de poziția lor în raport cu gradientul de concentrație al acelei molecule date generat de celula de semnalizare. Molecula responsabilă de diferențiere în acest caz se numește morfogen.
Tipuri de raportare
Urmează cinci căi posibile de comunicare celulară: autocrină, dependentă de contact, paracrină, endocrină și sinaptică.
Comunicarea autocrină apare atunci când celula țintă este aceeași celulă de semnalizare. Este o strategie populară pentru unele celule canceroase.
Comunicarea dependentă de contact apare atunci când moleculele de semnalizare se găsesc pe suprafața unei celule și semnalează celulele țintă prin legarea la ele fără a părăsi-o. Este utilizat pe scară largă de unele tipuri de leucocite, cum ar fi limfocitele.
Comunicarea paracrină apare atunci când molecula de semnalizare acționează la o distanță foarte mică asupra diferitelor celule țintă plasate lângă celula de semnalizare. Viteza acestei semnalizări depinde de distanța celulelor țintă și de răspunsul generat care poate apărea în fracțiuni de secundă, dacă are ca scop schimbarea conformației unei proteine, dar poate dura și minute sau ore când schimbă gena expresie. O variantă a acesteia poate fi considerată comunicarea prin joncțiunile gap, realizată cu molecule mici care difuzează prin conexiunile care leagă citoplasma a două celule adiacente. De asemenea, permit uniformizarea condițiilor electrice ale celulelor pe care le interconectează.
Comunicarea endocrină apare atunci când molecula de semnalizare, adesea transportată în sânge, acționează asupra unei celule țintă îndepărtate. Această moleculă este adesea un hormon. Este un tip de semnalizare relativ lent, a cărui viteză depinde de fluxul sanguin sau de fluidul în care este conținută molecula (o alternativă ar putea fi lichidul cefalorahidian). Concentrația unui hormon respectă ceea ce s-a spus despre concentrațiile moleculelor de semnalizare, stabilindu-se de obicei în jurul valorii de 10-8 M sau mai mici.
Neuronii folosesc un anumit tip de semnalizare care constă în producerea de potențiale de acțiune care rulează de-a lungul axonilor lor până când ajung la structuri specializate numite sinapse , care conțin vezicule în care sunt depozitați neurotransmițători; ca răspuns la un potențial de acțiune, unele dintre aceste vezicule se fuzionează cu membrana presinaptică, eliberează neurotransmițători în fanta sinaptică și aceștia se leagă de receptori specifici localizați pe membrana postsinaptică, declanșând în cele din urmă un potențial de acțiune suplimentar sau, în orice caz, un răspuns pe neuron sau pe celula țintă și apoi să fie degradat de enzime specifice (de exemplu acetilcolinesteraza). Tipul de comunicare descris, cel mai frecvent în sistemul nervos, este sinapsa chimică, dar există și sinapse electrice. Comunicarea sinaptică este foarte rapidă, deoarece potențialul de acțiune poate atinge viteze de 100 m s, iar celula țintă este separată de butonul sinaptic numai de fanta sinaptică îngustă, de câteva zeci de nanometri lățime. Trecerea de la membrana presinaptică la membrana postsinaptică este foarte rapidă, în ordinea unei milisecunde. Concentrația neurotransmițătorilor în fanta sinaptică este mult mai mare decât cea a altor molecule de semnalizare și poate depăși 10 4 M, în medie de zece mii de ori mai mult decât un hormon, deși scade rapid în urma degradării de către enzime specifice. De asemenea, este adevărat că receptorii pentru neurotransmițători au, spre deosebire de cei hormonali, o afinitate mult mai mică și tocmai această caracteristică garantează detașarea promptă și degradarea rapidă (sau recuperarea de către unele proteine) a neurotransmițătorului de semnalizare.
Receptorii
Receptorii sunt proteine care leagă molecule specifice. Specializarea unei celule determină ce tipuri de receptori sunt exprimați și care nu sunt exprimați. Un receptor poate ajuta la determinarea răspunsului celular la o moleculă de semnalizare pe baza stării sale de activare. De fapt, majoritatea receptorilor nu sunt pur și simplu activi sau inactivi, ci pot lua conformații care îi fac parțial activi chiar și cu diferite niveluri posibile de activare. Unele proteine, de exemplu, nu sunt activate complet după o singură fosforilare, dar necesită două fosforilări sau două fosforilări și interacțiune cu subunitatea unei a doua proteine și așa mai departe. Se disting receptorii de suprafață și receptorii nucleari.
Receptorii de suprafață leagă molecule de semnalizare care nu ar putea traversa membrana plasmatică altfel sunt hidrofile (la fel ca unii neurotransmițători) și sunt cel mai frecvent tip de receptori. Pot fi proteine extrinseci sau proteoglicani, dar cel mai frecvent sunt proteine transmembranare. Se disting trei clase mari de receptori de suprafață: receptorii conectați la canalele ionice, receptorii conectați la proteinele G și receptorii conectați la enzime. În plus față de aceste trei clase, există și alți receptori de suprafață care nu sunt asociați cu niciunul dintre cei trei.
Receptorii conectați la canalele ionice se găsesc în toate structurile specializate în transmiterea semnalului prin sinapse și în special pe membranele presinaptice și postsinaptice. Liganzii lor sunt neurotransmițătorii care deschid sau închid canalele ionice cu care este asociat fiecare dintre acești receptori, rezultând un potențial de acțiune asupra celulei țintă sau un alt tip de răspuns. Sunt proteine transmembranare cu mai multe treceri.
Receptorii legați de proteinele G au primit odată ligandul acționând prin activarea proteinelor G (adică a unei proteine trimerice care leagă GTP) adiacente acestora, care la rândul lor determină reglarea activității unei proteine țintă legată în general de membrană, dar separată de receptor . Activitatea proteinei țintă poate fi foarte diferită, poate fi un canal ionic, precum și o enzimă implicată într-o cale metabolică. De obicei sunt proteine transmembranare cu șapte treceri formate din helice α.
Receptorii legați de enzime sunt proteine transmembranare cu o singură trecere care pot acționa direct ca enzime sau pot activa alte enzime, de obicei prin fosforilare (mulți sunt, prin urmare, proteine kinaze ). În primul caz, situsul lor catalitic este intracelular, în timp ce domeniul de atac al ligandului este extracelular.
Receptorii nucleari leagă moleculele lipofile care pot traversa liber membrana plasmatică și ulterior se leagă de aceasta; în acest din urmă caz ele sunt de obicei proteine care reglează genele. Sunt proteine intrinseci sau mai frecvent proteine citosolice sau nucleare. Toți receptorii nucleari sunt legați între ei atât de mult încât aparțin unei singure familii, superfamilia receptorilor nucleari. Un receptor nuclear este format în ordine de un domeniu de activare a transcripției, situat lângă capătul N-terminal, un domeniu de legare a ADN-ului, apoi un domeniu de atașare a ligandului, care este plasat în schimb lângă capătul C-terminal. Când ligandul atacă, domeniul de atac al ligandului tinde să se închidă în sine pentru a prinde molecula. Aproape toți receptorii nucleari sunt formați printr-o alternanță de helice α și lanțuri intermediare fără structură caracteristică. Receptorii nucleari sunt inactivi înainte de atacul ligandului lor, deoarece sunt legați de proteinele inhibitoare. Când primesc molecula adecvată își schimbă conformația și atrag proteine activatoare care le activează. În acest moment, aceștia se pot deplasa din citoplasmă în nucleu sau își pot începe direct acțiunea (reglarea genei, activarea sau inactivarea transcripției). Genele transcrise direct de receptorul nuclear determină răspunsul primar în timp ce proteinele produse de aceste gene activează sau inhibă alte gene care constituie răspunsul secundar, sau proteinele răspunsului primar pot inhiba transcrierea propriilor gene (feedback negativ). Timpii pentru ca acest lucru să se întâmple sunt variabile, între cele două răspunsuri pot trece minute sau ore.
Al doilea mesager și proteinele de semnalizare intracelulare
Mesagerii secundari sau mediatorii intracelulari sunt molecule mici de semnalizare intracelulare generate ca răspuns la activarea receptorilor de către ligandul lor și proteinele la care sunt cuplați. Acestea sunt generate în număr mare de unele enzime specifice (cum ar fi adenilat ciclaza, guanilat ciclaza) de pe substraturi (cum ar fi ATP, GTP, PIP 2 ) și pot fi atât hidrofile (cum ar fi AMPc, cGMP, Ca 2+ ), cât și hidrofobe ( precum diacilglicerolul). În acest fel, semnalul este amplificat, astfel încât sute sau mii de molecule mesager secundare sunt generate pentru o singură moleculă de ligand legată de receptor. Al doilea mesager se leagă de proteinele de semnalizare intracelulare care constituie în general o cascadă de proteine de semnalizare sau o rețea. Aceste proteine pot acționa ca semnalizatori pentru proteina de semnalizare ulterioară, ca proteine de schelă pentru complexul de semnalizare (în acest caz aduc două sau mai multe proteine succesive împreună, facilitând interacțiunea lor), ca transductori de semnal, ca amplificatori de semnal, ca integratori ai semnalului , de la difuzoarele de semnal către alte căi de semnalizare, de la transportatorii acelorași proteine către o destinație specifică din celulă sau de la regulatorii altor proteine de semnal. În cele din urmă o cascadă de semnalizare activează una sau mai multe proteine efectoare.
Majoritatea proteinelor de semnalizare folosesc fosforilarea sau asocierea cu GTP pentru a acționa ca întrerupătoare și, prin urmare, își asumă o stare activă și una inactivă. În cazul reglării prin fosforilare, o proteină X este activată de o protein kinază specifică care transformă ATP în ADP pentru a atașa o grupare fosfat la proteina X generică, care va fi ulterior dezactivată de o proteină fosfatază care va elimina gruparea fosfat. Proteina kinază nu activează întotdeauna proteina X, uneori defosforilarea se activează de fapt. Protein kinazele sunt în general fie serină / treonin kinază (deci fosforilează proteina X pe serină sau treonină) sau tirozin kinază sau alte tipuri mai puțin frecvente. Fosfatazele proteice tind, de obicei, să fie mai puțin specifice în ceea ce privește proteinele asupra cărora acționează decât protein kinazele, după ce toți oamenii au 250 de gene protein-kinază și doar 150 gene proteice fosfatazice. Aproximativ 30% din proteinele umane suferă fosforilare. În cazul reglării de către proteinele G, starea activată a proteinei este determinată atunci când aceasta leagă GTP și starea inactivă când leagă PIB. Nu există enzime care să convertească GTP legat în PIB, deoarece activitatea GTPazei este intrinsecă proteinei G. Cu toate acestea, există și GTPaze monomerice care sunt inactive atunci când PIB este legat de acestea și active când GTP este legat. Aceste proteine, spre deosebire de proteinele G, sunt monomerice și non-trimerice și necesită un factor de schimb nucleotidic guanilic (GEF) pentru înlocuirea PIB cu GTP (activare) și o proteină care activează GTPaza (GAP) care transformă GTP în PIB cu fosfat eliberare (inactivare).
Alte proteine sunt activate sau inactivate prin ubiquitinare, proteoliză sau prin legarea la anumite molecule. Unele proteine de semnalizare intracelulare acționează ca integratori de semnale de la căi de semnalizare diferite și sunt activate numai dacă primesc semnale de la acele căi date. Proteinele de schelă, pe de altă parte, pot fi reduse la trei mari categorii. Ele pot fi proteine specifice schelelor prezente în citoplasmă care se apropie de receptori atunci când începe semnalizarea și în același timp atrag proteinele complexului de semnalizare din care fac parte, aducându-le mai aproape și facilitând posibilitatea interacțiunii, accelerând astfel semnalizarea în sine. Același receptor se poate autofosforila în poziții multiple și poate acționa ca o proteină de schelă. În cele din urmă, unele fosfolipide speciale pot fi fosforilate în continuare și pot servi ca situri de andocare pentru unele proteine intracelulare. Interacțiunea dintre proteinele de semnalizare intracelulară este facilitată de domeniile proteice comune multora dintre ele, care sunt interschimbabile și se pot lega în numeroase situri de orice proteină fără a-i modifica plierea. Printre cele mai frecvente sunt SH2 și SH3 (domeniile de omologie Src2 și Src3), domeniile de omologie la pleckstrină (PH), domeniile de legare la fosfotirozină (PTB).
Reglarea semnalului
Cele două modalități principale prin care o celulă își reglează proteinele de semnalizare sunt feedback-ul negativ și feedback-ul pozitiv.
În feedback negativ, un semnal de ieșire își inhibă propria producție. Feedback-ul negativ scurtează și limitează nivelul de răspuns și face sistemul de semnalizare mai puțin expus la posibile perturbații. În unele căi de semnalizare, feedback-ul negativ acționează cu o întârziere considerabilă, provocând răspunsuri oscilatorii atâta timp cât stimulul adecvat este prezent, în timp ce dacă întârzierea este scurtă, se determină răspunsuri de adaptare rapide și intense.
În feedback pozitiv, un semnal de ieșire favorizează producția sa, amplificându-l. În multe cazuri feedback-ul pozitiv acționează favorizând moderat propria producție, dar în unele cazuri îl mărește puternic, într-un mecanism similar cu „totul sau nimic”, atât de mult încât enzima, dincolo de un anumit prag, devine brusc mult mai activ și rămâne moderat activ chiar și atunci când semnalul a încetat. În acest caz vorbim despre un sistem instabil.
Când o celulă este expusă continuu la un semnal generat de o moleculă care acționează ca un stimul, apare fenomenul de adaptare sau desensibilizare, care funcționează ca un feedback negativ cu o scurtă întârziere. Desensibilizarea poate apărea prin sechestrarea complexului receptor + ligand din endosomii din citoplasmă, în acest caz în general ligandul rămâne în endozomi și este ulterior degradat în timp ce receptorul este eliberat și revine la membrana plasmatică sau prin reglarea descendentă a receptor, în acest caz complexul receptor + ligand este internat într-un endozom care fuzionează cu un lizozom care degradează atât ligandul, cât și receptorul. Receptorul poate fi totuși inactivat de o moleculă intracelulară care îl face incapabil să se lege de ligandul normal dacă concentrația extracelulară este prea mare sau dacă o proteină funcțională de receptor poate fi inactivată sau o proteină care inhibă receptorul poate fi inactivată interacțiunea receptorului cu proteinele din aval.
Semnalizare prin receptorii legați de proteina G
Receptorii legați de proteina G sau GPCR ( receptori cuplați cu proteina g ) sunt omniprezente în toate eucariotele și constituie cea mai mare familie de receptori de suprafață, cu aproximativ 700 de gene la om. Liganzii lor au o natură foarte variabilă, pot fi peptide, neurotransmițători, hormoni, aminoacizi și derivații lor, acizi grași și derivații lor, proteine și fotoni, fiecare se poate lega la un singur GPCR, dar și la numeroși GPCR, în acest caz frecvent aceste GPCR se găsesc în diferite celule. Nu toți liganzii pentru GPCR sunt cunoscuți pentru care unii receptori sunt orfani. În ciuda numărului lor, toate sunt alcătuite din proteine transmembranare cu șapte treceri, cu un domeniu extracelular în care ligandul se leagă (de dimensiuni diferite în funcție de ligand) și sunt cuplați cu proteinele G pe care le folosesc pentru a transmite semnalul în interiorul celulă.
Simțurile vederii, mirosului și gustului sunt mediate de receptori de acest tip. GPCR-urile sunt desensibilizate prin inactivarea receptorului, astfel încât nu mai pot interacționa cu proteinele G, prin sechestrarea receptorului care este interiorizat în endozomi sau prin reglare descendentă, care implică internalizarea și distrugerea receptorului și a ligandului. Toate aceste trei procese depind de fosforilarea serinei sau treoninei specifice GPCR de către PKA sau PKC sau de o kinază GPCR (GRK). Odată ce fosforilarea este efectuată, receptorul este complet inactivat prin legarea la o proteină din familia de arestine, care previne legarea de proteinele G și cuplează receptorul cu gropile acoperite cu clatrin, favorizând internalizarea lor care poate duce la degradarea receptorului sau temporar sechestrarea și eliberarea ulterioară și reintegrarea în membrana plasmatică.
Fiecare GPCR este cuplat la o proteină G care este fie legată de fața citoplasmatică a membranei plasmatice lângă receptor, fie atașată la receptorul însuși chiar și atunci când ligandul nu este legat și, prin urmare, receptorul este inactiv. Când este legată de membrană, o fac prin ancore lipidice care se ramifică de la subunitățile α și γ. Proteinele G sunt numeroase, dar toate au aceeași structură generală. Sunt proteine trimerice compuse dintr-o subunitate α, β și γ. Subunitatea α se leagă în starea inactivă PIB și GTP în starea activă. Când ligandul se leagă de un GPCR acționează ca o proteină GEF pentru proteina G la care este cuplat, promovând astfel schimbul de PIB cu GTP pe subunitatea α, activându-l și determinându-i să-și schimbe conformația. În majoritatea cazurilor, dar nu întotdeauna, la activare, proteina G se împarte în subunitatea α și într-un complex format din subunitățile β și γ. Proteina G nu are nevoie de GAP pentru a transforma GTP în PIB și pentru a se dezactiva, dar o face în mod spontan, deoarece subunitatea α are activitate GTPază, cu un timp variabil în funcție de capacitatea sa de hidrolizare. De obicei, timpul este scurt deoarece această subunitate acționează asupra proteinei sale țintă sau asupra unui regulator de semnalizare a proteinei G (RGS) care acționează ca GAP. Există aproximativ 25 de proteine RGS, fiecare dintre ele acționând asupra unui grup specific de proteine G.
Prin cAMP - PKA
Subunitatea α a proteinei G interacționează cu adenilat ciclaza (o enzimă transmembranară cu situsul activ intracelular) activând-o, aceasta la rândul său transformă ATP din citoplasmă în AMP ciclic (AMPc), unul dintre cei mai comuni mesageri secundari ai celula. amplificarea semnalului, deoarece atacul unui ligand determină sinteza a mii de molecule mesagere secundare în câteva secunde. Concentrația de AMPc în celulă variază , astfel , de la 10 -7 M în condiții normale până la 2,10 -6 M. adenilat ciclaza este la rândul său , reglată de o proteină G de stimulare (G s), care poate fi proteina G menționată mai sus., și o proteină G inhibitoare (G i ). CAMP se leagă de protein kinaza cAMP-dependentă (PKA). Odată ce semnalizarea este completă, AMPc este transformat rapid de AMPc fosfodiesterază în adenozină-5'-monofosfat (5'-AMP). PKA este un tetramer compus din două subunități reglatoare, de care se leagă două molecule AMPc pentru fiecare subunitate și două subunități catalitice.
Legarea implică disocierea proteinei în dimerul de reglare și în cele două subunități catalitice (care acționează ca monomeri), care sunt activate. Astfel de subunități fosforilează serine sau treonine dintr-o gamă largă de proteine țintă care pot fi alte proteine de semnalizare precum și proteine efectoare. Există proteine de ancorare ale kinazei A (AKAP) care se leagă de cele două subunități reglatoare ale PKA și le leagă de o componentă a citoscheletului sau de membrana unui organet, facilitând astfel localizarea unei părți a PKA intracelulară. În ciuda numelui lor, AKAP-urile nu leagă exclusiv PKA, ci și alte proteine, în acest fel le pot face să interacționeze cu același PKA, un exemplu este aceeași fosfodiesterază AMPc care poate astfel hidroliza cu ușurință cele patru molecule AMPc prezente pe subunitățile de reglare. Mai mult, această proteină este activată prin fosforilare de către PKA în sine, care, prin urmare, reglează activarea acesteia prin feedback negativ.
În unele celule, PKA poate activa genele care codifică anumiți hormoni, cum ar fi somatostatina, deoarece, după intrarea în nucleu, fosforilează proteina CREB ( proteina de legare CRE ), activând-o, care la rândul său se leagă de secvențele cunoscute sub numele de CRE ( element de răspuns AMPc ) plasat în regiunea de reglare a genei respective. În acest moment, o a doua proteină, coactivatorul transcripțional CBP ( proteina de legare CREB ) se leagă de CREB și complexul activează transcrierea genei. La fel ca în toate căile care implică transcrierea genelor, timpul necesar în acest caz nu este de câteva minute, deoarece ar putea fi dacă PKA activează alte proteine de semnalizare, ci ore. Se crede că acest proces este asociat cu formele de învățare și de memorie ale creierului. În alte celule, AMPc nu acționează asupra PKA, ci ca un ligand activator, de exemplu pentru canalele ionice ale membranei plasmatice (ca în neuronii olfactivi). De asemenea, poate activa Rap1, o GTPază monomerică care promovează aderența celulelor mediată de integrină.
Prin PIP2 - fosfolipază C.
Unele proteine G (numite G q ) nu activează adenilat ciclaza sau guanilat ciclaza, ci fosfolipaza C-β (PLCβ), de exemplu ca răspuns la legarea cu vasopresina în ficat și acetilcolina în pancreas și mușchiul neted. Fosfolipaza C-β este o enzimă care acționează asupra fosfatidilinozitol-4,5-difosfatului (PIP 2 ), un fosfolipid prezent în stratul interior al membranei plasmatice, transformându-l în inozitol-1,4,5-trifosfat (IP 3 ) și în diacilglicerol (DAG). IP 3 este solubil în apă și se difuzează în citosol prin legarea la canalele de eliberare Ca 2+ reglementate de IP 3 sau pur și simplu la receptorii IP 3 încorporați în membrana reticulului endoplasmatic neted (REL). Legarea IP 3 cu aceste canale induce scurgerea de Ca 2+ din reticulul endoplasmatic neted, crescând rapid concentrația sa în citosol. Ulterior, excesul de ioni Ca 2+ este returnat reticulului endoplasmatic neted de canalele Ca 2+ controlate de depozit. IP 3 la sfârșitul semnalizării este degradat de fosfatază la IP 2 sau fosforilat de kinază pentru a forma IP 4 , care la rândul său poate acționa ca o moleculă de semnalizare. Între timp, DAG poate fi utilizat pentru a forma acid arahidonic sau eicosanoizi, cum ar fi prostaglandinele, dar cel mai frecvent se lipeste de membrana plasmatică și activează protein kinaza dependentă de Ca 2+ (PKC), o serin-treonin kinază.
PKC este activat atât de DAG, cât și de Ca2 + eliberat de IP 3 , apoi fosforilează proteinele țintă, care variază în funcție de celula în care se efectuează semnalizarea. Există o a doua clasă de PKC, numită PKC atipică, deoarece nu este activată de DAG și Ca 2+ . Excesul de Ca 2+ din citosol este pompat în afara celulei de pompe proteice speciale plasate pe membrana plasmatică care consumă ATP, deoarece acestea trebuie să pompeze împotriva gradientului, fiind concentrația de Ca 2+ extracelular de zece mii de ori mai mare decât intracelularul unul (10 -3 M vs 10 -7 M). Fără ele, folosind canale de calciu, Ca 2+ ar tinde să curgă în citoplasmă. În plus față de pompele Ca 2+ , neuronii mai posedă și antiporturi Ca 2+ / Na + . Alte pompe de calciu sunt prezente în mitocondrii. Il Ca 2+ in seguito a rilascio dai recettori leganti IP 3 si diffonde nel citoplasma sotto forma di onde oscillatorie della frequenza di pochi secondi con picchi chiamati spike fino a che la segnalazione non termina. I picchi parrebbero essere generati dal fatto che il Ca 2+ rilasciato dai recettori per IP 3 attiva anche i recettori rianodinici che liberano ulteriori ioni Ca 2+ nel citoplasma, attuando un feedback positivo.
Tuttavia, oltre una certa concentrazione, il Ca 2+ inibisce un suo ulteriore rilascio da tali canali. In alternativa all'attivazione di PKC il Ca 2+ può attivare la calmodulina, una proteina molto abbondante nel citoplasma di tutte le cellule (ne costituisce sino all'1%). La calmodulina è una proteina dalla forma a manubrio, formata da due estremità globulari leganti ciascuna due ioni Ca 2+ interconnesse da un α-elica. La calmodulina inizia ad attivarsi con due ioni Ca 2+ legati, cambiando la sua conformazione, ma è completamente attiva solo con quattro ioni calcio. La calmodulina non possiede attività enzimatica e si lega ad altre proteine della cellula attivandole; ne sono esempi la pompa del Ca 2+ posta sulla membrana plasmatica che, come detto, riequilibra la concentrazione di ioni Ca 2+ nel citoplasma. Un bersaglio molto comune del complesso Ca 2+ -calmodulina sono le chinasi dipendenti da Ca 2+ -calmodulina (CaM-chinasi). Tali proteine spesso fosforilano fattori di trascrizione, attivandoli o inattivandoli. La CaM-chinasi II presente nei neuroni sembra essere fondamentale in alcuni processi di memoria cellulare, viene infatti inizialmente attivata da Ca 2+ -calmodulina, ma successivamente si autofosforila su residui di serina e treonina e resta attiva anche quando il Ca 2+ è stato sottratto finché non viene spenta da alcune fosfatasi.
Segnalazione tramite recettori collegati ad enzimi
I recettori collegati ad enzimi sono proteine transmembrana, generalmente a singolo passaggio, che possiedono un dominio extracellulare per il legame con il ligando e un dominio citosolico che possiede attività enzimatica intrinseca o è associato a diverse proteine, tra cui enzimi. Malgrado la diversità strutturale, molti recettori collegati ad enzimi attivano le stesse vie di segnalazione dei GPCR . Se ne distinguono sei classi.
I recettori tirosina chinasi o RTK sono proteine con un dominio citosolico capace di autofosforilarsi su specifici residui di tirosina , che possono fungere da ancoraggio per altre proteine citosoliche, inoltre possono fosforilare altre specifiche proteine.
I recettori associati a tirosina chinasi sono proteine con dominio citosolico privo di attività autofosforilante, ma attivano le loro vie di segnalazione reclutando tirosina chinasi citoplasmatiche.
I recettori serina/treonina chinasi sono proteine con dominio citosolico capace di autofosforilarsi su specifici residui di serina o treonina , ma possono fosforilare gli stessi amminoacidi anche su proteine che sono loro associate.
I recettori associati a istidina chinasi attivano un' istidina chinasi che si autofosforila su residui di istidina e poi trasferisce un gruppo fosfato ad una seconda proteina di segnalazione.
I recettori guanilato ciclasi possiedono un'attività enzimatica che permette loro di produrre direttamente cGMP da GTP, agendo quindi come una guanilato ciclasi .
Le tirosina fosfatasi simili a recettori possiedono un'attività fosfatasica che rimuove il fosfato da istidine su alcune proteine di segnalazione. I ligandi di queste proteine non sono conosciuti e sono perciò considerati recettori orfani. Talvolta non vengono neppure considerati recettori.
Recettori tirosin-chinasici
I recettori tirosin-chinasici (RTK) sono una delle tipologie più comuni di recettori di superficie collegati agli enzimi. I loro ligandi spesso sono fattori di crescita o neurotrofine come VEGF , NGF , EGF , HGF , FGF, MCSF, PDGF, IGF-1 ei recettori assumono spesso il nome dal loro ligando. Una classe di otto proteine extracellulari dette efrine (correlate con la migrazione e la crescita assonica) è strettamente associata ad alcuni RTK chiamati recettori Eph (dall'inglese ephrine ), codificati da 13 geni sui 60 geni umani per RTK . Le efrine quando si legano al loro recettore Eph attivano sia se stesse che il recettore, determinando cambiamenti sia nella cellula di segnalazione che nella cellula bersaglio. In generale un recettore RTK in forma inattiva è costituito da due monomeri a singolo passaggio che vengono attivati e dimerizzati una volta che il ligando si lega al dominio extracellulare. La dimerizzazione comporta la transautofosforilazione (un monomero fosforila l'altro e viceversa) su residui specifici di tirosina. Talvolta, come per il recettore dell'insulina, l' RTK in forma attiva è un tetramero. Le tirosina fosforilate aumentano l'attività chinasica dell' RTK e fungono da siti di legame per proteine intracellulari specifiche per ciascuna tirosina.
Il legame avviene perché tali proteine sono in grado di riconoscere la tirosina fosforilata e la conformazione dell' RTK attorno ad essa. Queste proteine sono attivate dal legame con la fosfotirosina e talvolta possono a loro volta autofosforilarsi o essere fosforilate su tirosine (questo secondo caso accade a IRS1, proteina associata al recettore per l'insulina). Tre delle più note proteine associate a RTK sono la fosfolipasi C-γ, che attiva la via dell'inositolo trifosfato e quindi PKC, la proteina Src che è a sua volta una tirosina chinasi e la fosfoinositide-3'-chinasi (PI 3-chinasi) che fosforila alcuni lipidi trasformandoli in siti d'attracco per altre proteine di segnalazione. Un dominio comune alle tre proteine elencate e con cui queste si legano alle fosfotirosine di RTK è SH2 ( Src homology 2 domain ), ma vi si può legare anche il dominio PTB ( phosphotyrosine binding domain ). Generalmente il legame di proteine alle fosfotirosine attiva una via di segnalazione, ma in alcuni casi la inibisce, come fa la proteina c-Cbl che monoubiquitina alcuni recettori segnalandoli per la degradazione (down-regolazione) che viene effettuata da proteine che contengono UIM (motivi d'interazione con l'ubiquitina) le quali dirigono il recettore verso vescicole rivestite da clatrina che infine si fondono con lisosomi. Fino a quando un recettore non è degradato può potenzialmente attivare ancora vie di segnalazione nell'endosoma.
Le proteine associate a RTK sono a loro volta spesso associate ad una proteina adattatrice, Grb2, tramite un dominio SH2. Grb2 possiede inoltre altri due domini SH3 per l'interazione con altre proteine. Uno dei domini SH3 spesso interagisce con Sos, una Ras-GEF (fattore di scambio guanilico per Ras ) adesa alla membrana plasmatica tramite un dominio PH ( pleckstrin homology domain ) che si lega ad un fosfoinositide trifosfato. Sos attiva Ras , una GTPasi monomerica che ha dunque GDP legato quando inattiva, per cui sostituisce GDP con GTP, a sua volta Ras attiva ulteriormente Sos in un circuito a feedback positivo. Successivamente fosfatasi specifiche rimuovono il fosfato da proteine a monte di Ras , inattivandole, mentre Ras stessa è inattivata da una Ras-GAP che idrolizza il GTP in GDP con liberazione di fosfato inorganico. Ras , quando attiva, fosforila il modulo della protein-chinasi attivata da mitogeni, comunemente noto come modulo della MAP chinasi. Ras attiva Raf (la chinasi MAPKKK), la quale a sua volta, con un meccanismo a cascata, attiva tramite fosforilazione Mek (MAPKK) che a sua volta attiva per fosforilazione Erk (MAPK). Erk entra nel nucleo e fosforila alcune proteine regolatrici di geni oppure attiva altre proteine. Tra i geni attivati ve ne sono alcuni che regolano la proliferazione cellulare, come le cicline, perciò non sorprende che mutazioni iperattivanti di Ras inducano la sovraespressione di queste proteine con conseguente iperproliferazione cellulare e dunque cancro. Ras infatti è mutata nel 30% delle neoplasie umane, il che ne fa uno dei bersagli preferiti delle cellule tumorali dopo p53 .
Il modulo MAP chinasi viene spesso spento dalla stessa Erk che inattiva Raf tramite feedback negativo, altre volte invece sono delle fosfatasi a doppia specificità (sia per tirosina che per treonina) ad inattivare il complesso rimuovendo i gruppi fosfato. Generalmente la cascata delle MAP chinasi funziona efficientemente grazie a proteine impalcatura che legano tutte le chinasi coinvolte migliorandone l'interazione a causa della prossimità indotta. In un'altra importante via (via PI3-chinasi-Akt) attivata dagli RTK una classe di proteine nota come PI 3 chinasi, attaccata normalmente alla coda di RTK , fosforila alcuni inositolo fosfolipidi (PIP 2 ) in siti multipli generando fosfoinositidi (PIP 3 ). I fosfoinositidi sono siti d'attracco ideali per molte proteine. I GPCR possono attivare la stessa via, tuttavia agiscono sulla classe Ib delle PI 3 chinasi, mentre gli RTK sulla classe Ia.
Per far terminale il segnale alcune fosfatasi, in particolare le PTEN, defosforilano i fosfoinositidi determinando il distacco delle proteine che vi si legano. Mutazioni in PTEN aumentano il rischio di sviluppare alcuni tipi di cancro. Ai fosfoinositidi si attaccano due proteine, Akt (detta anche protein-chinasi B, PKB) e la protein-chinasi 1 dipendente da fosfoinositidi (PDK1). Il legame delle due proteine ad un fosfoinositide determina l'attivazione di Akt da parte di PDK1. Akt attiva una GAP chiamata Tsc2 che a sua volta disattiva Rheb. Rheb è una GTPasi attiva con GTP legato che attiva il complesso proteico mTOR. mTOR nei mammiferi è costituita dal complesso mTOR1, con legata la proteina raptor, oppure mTOR2, con legata la proteina rictor. Sono entrambe legate alla crescita e alla sopravvivenza cellulare ed interagiscono con la GTPasi monomerica Rho che a sua volta agisce sul citoscheletro. Akt inoltre, oltre ad attivare Tsc2, viene fosforilata da mTOR. A questo stato d'attivazione Akt è in grado di fosforilare i complessi Bad-proteina antiapoptotica (Bad è una proteina proapoptotica), determinando il distacco di Bad dalla proteina antiapoptotica e la sua inattivazione da parte della proteina 14-3-3. Il risultato netto è un'inibizione dell'apoptosi coerente con la proliferazione e la crescita cellulare.
Recettori associati a tirosina chinasi
I recettori associati a tirosina chinasi non possiedono attività tirosin-chinasica intrinseca ma sono associati covalentemente a tirosine chinasi sul lato citoplasmatico. La classe più comune di tirosin-chinasi associate a questo tipo di recettori è la famiglia delle Src, che comprende Src, Yes, Fgr, Fyn, Lck, Lyn, Hck e Blk. Ciascuna proteina Src contiene un dominio SH2 con cui si lega al recettore e almeno un dominio SH3 con cui interagisce con altre proteine. Proteine della famiglia Src sono attivate anche da proteine G e collegate a recettori GPCR . I recettori associati a tirosina chinasi comprendono alcuni recettori per l'antigene, per le interleuchine, per le integrine e per le citochine. Nel caso dei recettori per le citochine la proteina con attività tirosin-chinasica a cui sono associati è una JAK chinasi o chinasi Janus. Si conoscono quattro JAK chinasi, chiamate JAK1, JAK2, JAK3 e TYK2.
I recettori sono due monomeri separati quando inattivati, ma il legame del ligando li fa dimerizzare ed avvicina le due JAK chinasi che si fosforilano per transautofosforilazione su tirosine. Una volta attivate, fosforilano tirosine anche sul recettore per le citochine. Le fosfotirosine attirano proteine regolatrici di geni della famiglia STAT ( signal transducers and activators of transcription ) normalmente presenti nel citoplasma. Tali proteine contengono un dominio SH2 che si lega alla fosfotirosina del recettore, dopodiché vengono fosforilate dalle JAK ed attivate. Le STAT che si legano al recettore sono generalmente STAT1, STAT2 o STAT5. Le due STAT fosforilate si staccano dal recettore e formano un omodimero o un eterodimero tramite il dominio SH2, dopodiché migrano nel nucleo e si legano ad elementi di risposta alle citochine, favorendone la trascrizione dei geni corrispondenti. La via viene spenta da tirosina fosfatasi che possono anche essere a doppia specificità (rimuovono cioè gruppi fosfato anche da serina o treonina oltre che da tirosina).
Recettori serina/treonina chinasi
Sono proteine transmembrana a singolo passaggio con almeno un dominio serina/treonina-chinasi sul lato citoplasmatico della membrana, capaci di autofosforilazione. Si distinguono in recettori serina/treonina chinasi di tipo I e di tipo II, sono entrambi omodimeri e strutturalmente simili. I ligandi principali di questo tipo di recettori sono le BMP (bone morphogenic proteins) e la famiglia del fattore trasformante di crescita β (TGFβ). Quando il ligando si lega al recettore appropriato, questo si autofosforila nel dominio con attività chinasica su serine o treonine specifiche, attirando una classe di proteine nota come Smad ed attivandole. Qualora il ligando sia una proteina della famiglia di TGFβ, sono coinvolte Smad2 e Smad3, qualora invece sia una BMP sono coinvolte Smad1, Smad5 o Smad8. Le cinque Smad nominate sono dette Smad attivate da recettore (R-Smad). Una volta fosforilata, come già visto per le STAT, Smad si dissocia dal recettore e si associa con Smad4 detta anche co-Smad in quanto si lega alle R-Smad per formare un complesso.
Il complesso così formato si dirige nel nucleo dove si lega a sequenze specifiche che aiutano ad attivare o inibire alcuni geni. La via è spenta dall'endocitosi dipendente da caveole dei recettori che legano BMP o proteine della famiglia di TGFβ; successivamente questi sono ubiquitinati e quindi degradati. In realtà, l'endocitosi può anche attivare ulteriormente questi recettori se endocitati in endosomi precoci derivati da vescicole rivestite da clatrina. Una proteina chiamata SARA (Smad anchor for receptor activation) si lega ai recettori migliorando l'efficienza della fosforilazione di Smad. Nel nucleo il complesso Smad è defosforilato e poi trasportato nuovamente nel citoplasma dove può iniziare un nuovo ciclo.
I recettori inoltre possono anche essere inibiti da Smad-inibitrici (Smad6 e Smad7) la cui trascrizione è favorita dalle R-Smad, si attua così una regolazione a feedback negativo. Le Smad inibitrici competono con le R-Smad per il legame al recettore, saturandone il dominio di legame e quindi impedendo alle Smad leganti il recettore di intervenire. Possono inoltre attirare Smurf, un'ubiquitina ligasi che marca il recettore per la degradazione, o, ancora, possono reclutare proteine fosfatasi che defosforilano il recettore, inattivandolo e si legano a Smad4 competendo con le R-Smad.
Segnalazione tramite proteolisi
Una delle vie di segnalazione più dirette ed ampiamente utilizzate nel corpo umano e in molti animali è quella che fa uso del recettore Notch. Notch è una proteina transmembrana a singolo passaggio che interagisce con la proteina Delta, posta sulla membrana plasmatica di una seconda cellula. Quando Notch è attivata da Delta una proteasi taglia la coda di Notch ed essa, dopo una successiva proteolisi da parte della γ-secretasi, migra nel nucleo attivando la trascrizione di una serie di geni tramite una proteina che nello stato latente funge da repressione ma in seguito ad interazione con la coda di Notch diviene un attivatore. I geni che influenza sono molti, ma i principali sono quelli della famiglia Hes, che codificano per proteine regolatrici inibitorie verso altri geni. Questa via è coinvolta nel differenziamento in senso non-neuronale di una cellula, dal momento che permette il blocco dei geni che specializzerebbero la cellula in senso neuronale. Una volta effettuata la proteolisi, il recettore non può più essere riutilizzato.
Note
Voci correlate
Comunicarea celulară și biofotonii
Comunicarea celulară și biofotonii
Omul este o structură extrem de complexă alcătuită din trilioane de celule, ale căror funcţionalităţi nu le cunoaştem în întregime. Dacă această afirmaţie vi se pare una pretenţioasă, reţineţi că doar în secundele în care aţi citit aceste cuvinte, într-o singură celulă din corpul dumneavoastră au avut loc peste 100.000 de reacţii chimice.
Căutări şi răspunsuri
Dr. Fritz Albert Popp s-a născut la 19 mai 1938 şi este biofizician german, specializat în biologie celulară şi fizică cuantică. Împreună cu colegii săi de la Universitatea din Marburg a început să studieze emisiile de biofotoni. Sunt acele particule care transmit energie termică sau cuante de lumină. Acestea au ca sursă un corp viu, animal sau vegetal. El a stabilit că aceste radiaţii sunt extrem de slabe, imperceptibile pentru organele noastre. Nu trebuiesc confundate cu bioluminiscenţa, care este produsă de unele organisme vii (licurici, peşti, mucegaiuri, ciuperci) printr-un proces chimic de oxidare a luciferinei.
S.O.S. la nivel celular
Biofotonii sunt produşi prin reacţiile care au loc în „programul” nostru genetic, în acidul dezoxiribonucleic (ADN). Ei sunt energie coerentă, purtătoare de informaţii şi sunt legaţi de funcţionarea sistemelor noastre biologice. În anul 1974, dr. V.P. Kazanciev, împreună cu echipa sa de cercetare din Novosibirsk, a anunţat că a detectat o comunicare intercelulară prin intermediul biofotonilor. Până în 1980, el a efectuat peste 12.000 de experimente. Acesta a descoperit, printre altele, că celulele rănite sau bolnave emit o cantitate mai mare de biofotoni. Acest lucru a fost interpretat ca fiind un semnal de alarmă, un SOS, prin care se anunţă o primejdie pentru organismul respectiv.
Comunicare celulară permanentă, codificată
Acest schimb de informaţii seamănă cu transmisiile prin intermediul fibrelor optice. Acest fapt, i-a determinat pe cercetători să se întrebe dacă noi nu suntem cumva nişte holograme care funcţionează prin comenzi primite din altă parte. În mai 2012, dr. Serghei Maiburov, de la Institutul de Fizică Lebedev din Moscova, a afirmat că între celulele organismelor vii există o comunicare permanentă, efectuată printr-un cod secret pe bază de lumină, descifrat numai la nivel celular.
Experimental, el a constatat că icrele de peşte emit biofotoni care pot favoriza dezvoltarea altor ouă de peşte, aflate în alt loc. Fluxurile biofotonice sunt emise în „rafale”, la intervale periodice şi seamănă cu o transmisie de date binare. „Este un fel de comunicare digitală ca între calculatoare. Analiza radiaţiilor de biofotoni dintre diferitele culturi de icre a demonstrat că informaţiile sunt codificate la fel ca la un algoritm analogic”, a spus savantul.
O inginerie bizară
Studii recente au demonstrat că sistemul de comunicaţii biofotonice este independent de mecanismul care produce fotonii. Iniţial s-a crezut că acestea sunt incoerente. Mai târziu însă, s-a constatat că sunt asemănătoare impulsurilor de la un laser. Dacă icrele de peşte comunică între ele, există şi alte sisteme complexe, ca de pildă creierul uman, care emit semnale biofotonice?
Comunicarea neuronală
Se ştie că neuronii, acele celule adaptate la recepţionarea şi transmiterea de informaţii, comunică între ei. Aceștia au rol determinant în funcţionarea organismului uman. Transmisiile se realizează prin reacţia unor substanţe chimice precum serotonina, dopamina şi noradrenalina. Serotonina participă la producerea somnului, a depresiei, anxietăţii, a senzaţiei de vomă, dar şi la reglarea pulsului, tensiunii arteriale şi temperaturii.
Dopamina este un neurotransmiţător care transportă informaţiile între celulele creierului. Aceasta ne ajută să simţim plăcerea, dar şi durerea şi este cea care ne influenţează în luarea deciziilor. Noradrenalina este responsabilă de transmiterea informaţiilor spre muşchii netezi (ex. la intestine) şi glande. Dar nu toate informaţiile ajung la creier. Unele sunt oprite la măduva spinării sau la bulbul rahidian. Este un sistem nervos autonom, care funcţionează independent de creier, îl ajută şi este mai rapid decât acesta.
Cea mai puternică emisie de lumină
Acest sistem este răspunzător de respiraţie, digestie, bătăile inimii, excreţie, sete și reproducere. Este vorba despre acele activităţi pe care le facem fară să gândim, permanent. În mod curios, şi probabil deloc întâmplător, dintre toate substanţele chimice din corpul nostru acestea au cea mai puternică emisie de lumină. Interferează cu sistemul nervos central şi sunt participante la emisiunile de biofotoni. Altfel spus, pe lângă transmiterea de informaţii pe căi electrice şi chimice, propagarea semnalelor se face şi prin biofotoni.
Calcule ipotetice telegrafice
Încă nu ştim bine unde se formează biofotonii, ştim însă că ei transmit o parte din informaţiile necesare morfogenezei. Adică acelui proces de formare a structurii organismelor vegetale şi animale. Mai ştim că acest schimb de informaţii seamănă foarte bine cu procedura standard prin care computerele fac schimb de date între ele. La oameni, gândurile sunt energie, care este purtătoare de informaţii. Informaţiile sunt transmise şi de biofotoni. Acești biofotoni, circulă cu viteze mai mari decât cea a luminii.
Gândul are viteze superluminice. Eu pot ajunge cu gândul la Proxima Centauri, cea mai apropiată stea de sistemul nostru solar, instantaneu. Luminii îi trebuie 4,22 ani ca să străbată această distanţă. Toate celulele organismelor vii, inclusiv ale omului, emit biofotoni. Fără să ştim, suntem într-un neîntrerupt flux de informaţii, într-un ocean în care energii nebănuite se interferează influenţându-ne permanent şi ireversibil viaţa.
(Nu sunt voturi încă)
(Nu sunt voturi încă)
Adaugă un comentariu









